
Altersklassen im Sport
In den leichtathletischen Sportarten werden nach dem Hochleistungsalter, meist nach dem 35. Lebensjahr, erbrachte Leistungen nach Alter und Geschlecht getrennt bewertet. Die nationalen und internationalen Leichtathletikverbände haben nach Geburtsjahr die Altersklassen in 5-Jahres Schritten festgelegt. Die Altersklassen (AK) beginnen mit dem 35. Lebensjahr (Jahrgang) und enden derzeit bei AK 105.
Welche Ursachen der Leistungsabnahme im Seniorenalter werden gegenwärtig diskutiert?
Theorien zum Altern
Zur Alterung gibt es zahlreiche Theorien, welche im Einzelnen die Komplexität der Alterung nur teilweise beschreiben können (Rose et al. 2008). In Tab. 1 sind die wichtigsten Theorien zu möglichen Ursachen der Alterung aufgeführt.
Im Seniorenalter kommt es bei Abnahme der motorischen Aktivität sowie gleichbleibender Energiezufuhr zur Zunahme von Körperfett und zu einem Verlust der fettfreien Körpermasse in der Muskulatur. Die allmählich nachlassend körperliche Aktivität, auch bei Sporttreibenden, führt zur Verringerung des Energie- und Grundumsatzes und letztlich zur Abnahme der sportlichen Leistungsfähigkeit. Weiterhin haben komplexe hormonelle Veränderungen Einfluss auf zelluläre Alterungsprozesse in den Organen. Das betrifft besonders die veränderten Funktionen von Hypothalamus-Hypophysen-Gonadalachse, Hypothalamus-Hypophysen-Nebennierenrinden-Achse sowie die Hypothalamus-Hypophysen-Thyroid-Achse (Jones & Boelaert, 2015). Auch das Wachstumshormon (GH), der Faktor IGF, Insulin und Prolaktin sowie die Darmhormone, das Vitamin D und das Pre-Rezeptor-Hormon-System beeinflussen die Alterungsprozesse in den Geweben. Die hormonellen Einflüsse auf das Altern sind so bedeutsam, dass daraus eine eigene Alternstheorie erarbeitet wurde (Russell & Kahn, 2007).
Unabhängig von den Alterungstheorien wird nachfolgend aufgeführt zu welchen erstaunlichen Leistungen Seniorinnen und Senioren noch fähig sind. Auffallend ist, dass die meisten Bestleistungen in den Altersklassen (AK) erst in den letzten 5 bis 10 Jahren vollbracht wurden (Abb. 1). Das spricht dafür, dass auch die alternden Sportlerinnen und Sportler ein Bestreben nach Erreichen von Bestleistungen, im Vergleich zu gleichaltrigen Athleten haben.
Tab. 1. Auswahl von Theorien zur Ursache der Alterung des Menschen (dvs Band 28 © Edition Czwalina)
Alterstheorien | |
Schadstoff Theorien | Licastro et al. (2005). Shanley et a. (2009). Sancar, Lindsey-Boltz, Unsal-Kacmaz & Linn (2004). |
Telomerhypothese des Alterns | Hartley, Kim, Prowse, Weinrich, Hirsch et al., (1992). |
Zelluläres Altern (Seneszenz) | Gilka, Stoian, Atanasiu & Virgolici (2007). |
Apoptose und zelluläre Veränderungen | Pollack & Leeuwenburgh (2001). |
Entzündungsaltern | Franceschi et al. (2000). |
Evolutionstheorien des Alterns:Programmiertes Altern und klassische Evolutionstheorien des Alterns | Gredilla & Barja (2005). Hughes & Reynolds (2005), Rose et al. (2008), Ljubuncic & Reznick (2009), Russell & Kahn (2007) |
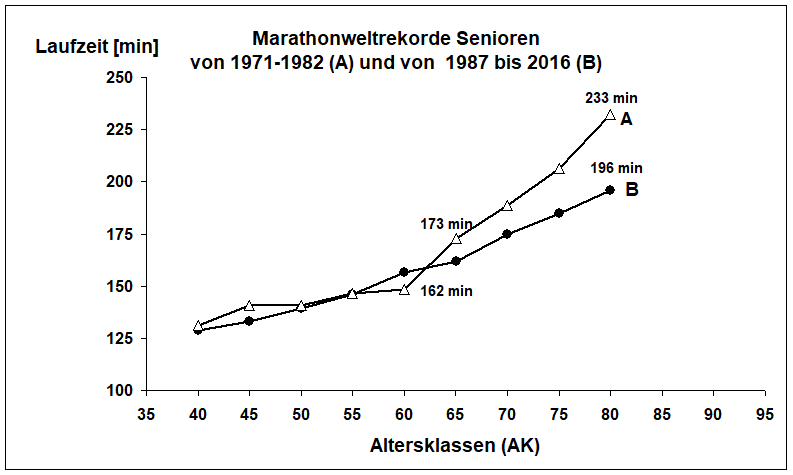
Abb. 1. Marathonweltrekorde Männer im Vergleich der Jahre 1971 bis 1972 und 1987 bis 2018. In den letzten 10 Jahren kam es zu einer deutlichen Leistungsverbesserung in der Laufzeit ab der AK 65.
Ursachen des Muskelschwundes im Alter
Sarkopenie
Die mit zunehmendem Alter abnehmende Muskelmasse wurde von Rosenberg (1997) erstmals als Sarkopenie bezeichnet. Die Sarkopenie ist nicht mit jenem Abbau der Muskelmasse bei krankheitsbedingter Inaktivität im jüngeren Alter identisch. Der physiologische Rückgang der Muskelmasse beruht auf einer Inaktivitätsatrophie, wie z. B. durch Ruhigstellung von Gliedmaßen bei Frakturen, krankheitsbedingtem Bewegungsmangel u. a.). Durch gezieltes Training ist die ruhiggestellte fettfreie Muskulatur (lean body mass) in wenigen Wochen wieder voll funktionsfähig.
Anders ist es bei der altersbedingten Sarkopenie. Hierbei wird die abnehmende Muskelmasse teilweise durch Bindegewebe und Fett ersetzt. Zwischen dem 50. bis 60. Lebensjahr nehmen die körpereigenen Fettdepots beim Normalbürger um etwa 2 kg zu (13-15 %). Ohne Training verminderte sich die aerobe Leistungsfähigkeit zwischen dem 30. und 70. Lebensjahr um 8-10 % in jeder Lebensdekade (Pollock, Lowenthal, Graves, & Caroll, 1993; Pollock, Mengelkoch, Graves, Lowenthal, Limacher et al., 1997).
Der Muskelschwund hat komplexe Ursachen; das Argument der Bewegungarmut reicht als alleinige Erklärung nicht aus. Der Verlust an Muskelfasern beginnt mit dem 50. Lebensjahr und führt bis zum 80. Lebensjahr zu einer Verminderung von 50 %. Damit erfolgt in jeder Lebensdekade eine durchschnittliche Abnahme an Muskelmasse um 15 %. Wenn diese Aussage für die Normalpopulation gelten sollte, so wäre denkbar, dass eine sportliche Aktivität eine Muskelmassenabnahme verhindern könnte. Mit zunehmendem Alter ist das aber nicht der Fall.
Das nachlassende Bedürfnis der Nahrungsaufnahme, welches unterschiedliche hormonelle Ursachen hat, fördert die Abnahme der Muskelmasse (Morley, 2001).


Der Verlust an kontraktiler Muskelmasse im Alter (Sarkopenie) erfolgt besonders für die schnell kontrahierenden ST-Fasern (Typ 2) (Lexell, 1995; Grimby, 1995; Power, Dalton & Rice, 2013). Ab dem 40. Lebensjahr erfolgt der Muskelschwund von 1-2 %/Jahr (Grimby, G., 1995). Das Ausmaß der Sarkopenie ist messbar an Kraftabnahme im Alter (Narici & Maffulli, 2010). Die Kraftverminderung betrifft beide Geschlechter.
Insgesamt sind die Ursachen des Muskelschwunds mit zunehmendem Alter, mit oder ohne Training, noch nicht vollständig geklärt (Roubenoff & Hughes, 2000). Am Abbau von Muskulatur scheinen mehrere Faktoren beteiligt zu sein und eine multifaktorielle Genese ist sehr wahrscheinlich. Zu den Veränderungen mit zunehmendem Alter zählen eine verminderte Steroidsynthese und eine abnehmende Synthese des Wachstumshormons sowie IGF1. Genetische Einflüsse auf die Alterung sind sehr wahrscheinlich. Hinzu kommen die zunehmende Bewegungsarmut sowie die damit zusammenhängende oder unabhängig davon verminderte Proteinsynthese. Der Einfluss abnehmender Proteinsyntheseraten nach sportlichen Belastungen wird subjektiv erlebbar durch die längeren Regenerationszeiten nach körperlicher Belastung. Zudem ist die Ernährung im Alter suboptimal, die Energieaufnahme ist vermindert. Die Zufuhr essenzieller Proteine nimmt ab. Bei trainierenden Senioren beiderlei Geschlechts ist der Muskelschwund geringer. Jedoch ist ab dem 80. Lebensjahr der Muskelschwund gleichgroß wie bei Untrainierten, er beträgt etwa 50 % der ursprünglichen Masse (Faulkner et al., 2007). Die drastischen Leistungsabfälle (Laufgeschwindigkeit) in den leichathletischen Disziplinen (z. B. Marathonlauf) weisen ab der AK 85 darauf hin (Abb. 2).
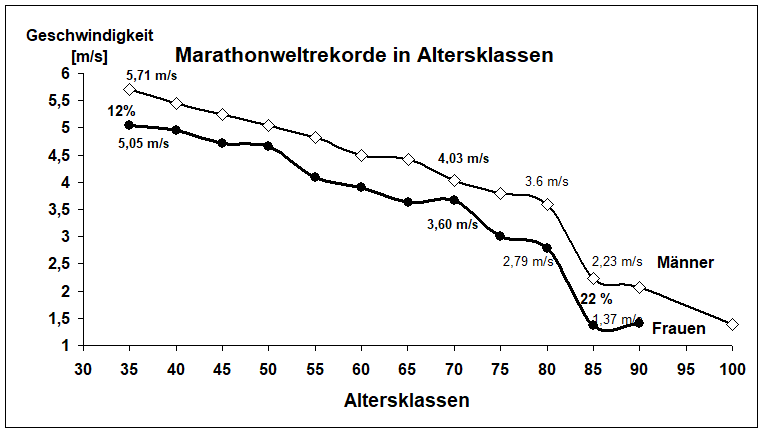
Abb. 2. Marathonweltrekorde (Laufgeschwindigkeit) von Männern und Frauen in den Alterklassen (AK) 35 bis AK 100. Stand 2019. Die Geschlechtsdifferenz in der Laufgeschwindigkeit nimmt von 12 % in der AK 35 auf 22 % in AK 85 zu. Nach der AK 80 ist der Abfall der Laufgeschwindigkeit bei Männern und Frauen drastisch.
Ein sportliches Training kann den Prozess des Muskelabbaus im Zeitraum der Alterung vermindern, aber nicht aufhalten.
Der Vergleich der Muskelmasse von Altersportlern mit 25jährigen Sportlern ist signifikant, d. h., die funktionelle Abnahme des neuromuskulären Systems im Seniorenalter ist trotz Training vorhanden, nur im verminderten Ausmaß (Power, Dalton & Rice, 2013). Je älter man wird, umso wahrscheinlicher ist die Sarkopenie, weil mit der Alterung auch die Entzündungsprozesse im Organismus zunehmen und diese bekanntlich auch als Modell einer Alterungstheorie gelten (s. Tab. 1). Mit zunehmendem Alter steigen bei motorischer Untätigkeit die entzündungsassoziierten Zytokine an, welche den Abbau der Muskelmasse fördern. Der Muskelschwund ab dem 40. Lebensjahr mit 1-2 %/Jahr ist im hohen Alter ein wesentlicher Grund für die Abnahme der Muskelkraft (Grimby, 1995). Diese Aussage wird z. B. durch eigene Analysen der kraftabhängigen Abnahmen von Weit- und Hochsprungleistungen im Seniorenalter bestätigt (Neumann, Hottenrott & Hottenrott, 2019). Die Schnelligkeit, beurteilt an der Geschwindigkeit beim 100-m-Lauf, nahm mit zunehmendem Alter langsamer ab als die erforderlichen Schnellkraftfähigkeiten im Hoch- und Weitsprung.
Muskel als sekretorisches Organ
Neu ist die Erkenntnis, den Muskel als sekretorisches Organ zu bezeichnen (Pedersen, 2013). Die Berechtigung wird davon abgeleitet, dass der Muskel hunderte Myokine produziert, welche eine Untergruppe der Zytokine sind. Die Zytokine sind spezielle Proteine, die das Wachstum und die Differenzierung von Zellen regulieren.
Die Muskulatur kann als sekretorisches Organ bezeichnet werden, da sie endokrine, parakrine und autokrine Botenstoffe freisetzt, die als „Myokine“ bezeichnet werden (Pedersen, 2013). Bisher sind über 600 Myokine, die als Untergruppe der Zytokine (Botenstoffe) gelten, beschrieben (Pedersen, 2011). Die Arbeitsgruppe um Pedersen (CIM Copenhagen, Dänemark) hat angeregt, dass Zytokine und andere Peptide, die von der Muskelzelle produziert, exprimiert und frei gesetzt werden als „Myokine“ zu klassifizieren (Pedersen, 2011 a). Das muskuläre Sekretom besteht aus mehreren Hundert sekretierten Peptiden (Pedersen, 2013). Die Muskelzelle kommuniziert bei Belastung mit anderen Organen. Die von den Muskelzellen exprimierten Myokine geben Signale an die Körperzellen ab, die zahlreiche Wirkungen entfalten können, so die Beeinflussung des Endothels, indem die Verzweigung von Blutgefäßen bei Kontraktion und Bewegung gesteigert wird (Pedersen, 2011). Das durch Belastung und Krafttraining erhöhte IL6 beeinflusst die Myogenese (Muskelfaserhypertrophie). Erst durch Bewegung (Sport) werden ausreichend Myokine gebildet, bei Bewegungsarmut sinkt die Bildung von Myokinen auf ein Minimum. Der Zustand der Bewegungsarmut im Alter kann ein Faktor dafür sein, dass die Kommunikation der unterbelasteten Muskulatur mit den Körperorganen nachlässt.
Die bei Muskelbelastung freigesetzten Myokine nehmen auch Einfluss auf zahlreiche Stoffwechselvorgänge und bewirken so die Vorbeugung von zahlreichen Krankheitsbildern (Pedersen, 2011; Pedersen & Febbraio, 2012).
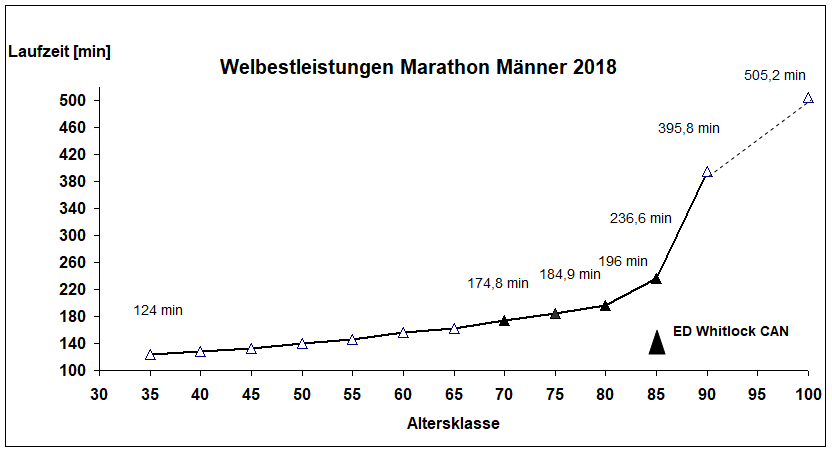
Abb. 3 Weltrekorde (Laufzeit) im Marathon Männer von 2018. Ab der AK 70 bis zur AK 85 bestimmte Ed Whitlock (Canada) die Weltbestzeiten im Marathon. Er lief mit 73 Jahren den Marathon noch in 2:54:48 h und die 10.000 m in 37:33 min.
Muskel und Telemeraseaktivität
Die Telomere sind die nicht kodierenden und einzelsträngigen Enden an den Chromosomen. Als Strukturelemente der DNA sind sie für deren Stabilität zuständig. Bei jeder Zellteilung verkürzen sich die Telomere. Wenn sie eine kritische Länge (4kbp) unterschreiten, kommt es zur Apoptose der Zellen (programmierter Zelltod). Das Enzym Telomerase kann diese Verkürzung wieder ausgleichen bzw. vermindern.
Die Telomerenerosion ist eine zentrale Komponente der Alterung (s. Tab. 1). Die in den Telomeren assoziierten Proteine regulieren die Seneszenz und das Überleben. Bei Ausdauertrainierten war eine gesteigerte Telomerasenaktivität nachweisbar. Beim Vergleich von 3 Trainingsgruppen, welche Ausdauer trainierten, ein intensives verkürztes Ausdauertraining durchführten (HIT) und nur Krafttraining absolvierten, gab es unterschiedliche Einflüsse auf die Telomerasenaktivität und Telomerenlänge.
Nur das Ausdauertraining aktivierte am stärksten die Telomeraseaktivität und erhöhte die Telomerenlänge an den Chromosomen (Werner at al., 2018). Das HIT-Training war vermindert wirksam und das Krafttraining zeigte keinen Einfluss auf die Telomereraseaktivität und Telomerasenlänge. Das Ausdauertraining reguliert die telomerstabilisierenden Proteine und schützt vor einer stressinduzierten Apoptose von Gefäßen.
Weitere Einflussfaktoren aus die Leistungsabnahme im Seniorenalter
Neben der Bewegungsarmut und dem Muskelschwund gibt es noch weitere Einflussfaktoren auf die Leistungsminderung, die sowohl bei Altersportlern als auch bei Normalpersonen wirken (Ryall, Schertzer & Lynch, 2008).
Mit zunehmendem Alter nimmt die Knochendichte ab, das betrifft besonders Frauen in der Menopause (Martyn-StJames & Caroll, 2008). Die im Alter abnehmende Proteinsynthese führt zur deutlichen Verlängerung der Erholungszeit nach körperlichen Belastungen. Demnach beruht die verminderte Belastbarkeit alternder Sportler auf der längeren Regenerations- bzw. Erholungszeit nach sportlichen Belastungen.
Die längere Regenerationszeit vermindert auch die Gesamtbelastbarkeit. Nach dem 50. Lebensjahr betrug die Trainings- und Wettkampfbelastung der untersuchten Altersläufer zwischen 1.000 bis 2.000 km/Jahr (Neumann, Hottenrott & Hottenrott, 2019).
Die maximale Sauerstoffaufnahme nimmt sowohl bei Untrainierten als auch bei trainierten Alterssportlern ab, nur bei letzteren erfolgt die Abnahme aufgrund der körperlichen Aktivität auf einem höherem Niveau bzw. erreicht nicht die Tiefwerte Untrainierter (Hawkins & Wiswell, 2003).
Da ältere männliche Sportler eine individuell unterschiedlich nachlassende Testostronbildung bzw. die Frauen eine nachlassende Androgenbildung mit Abnahme des Öestrogens aufweisen, folgt, dass bei beiden Geschlechtern eine abnehmende Proteinsynthesegeschwindigkeit in der Muskulatur stattfindet. Im Alterungsprozess kommt es zu komplexen Veränderungen im Hormonsystem. Die Abnahme des freien Testosterons beträgt nach dem 25. Lebensjahr 1,2 % jährlich und des Wachstumshormons (GH) 18 % ab dem 30. Jahr je Lebensdekade beim Mann. Auch das Östrogendefizit und der Progesteronmangel nach der Menopause verursachen bei der Frau (Vermeulen & Kaufmann 1995; Jockenhöfel, 1999) eine Leistungsabnahme. Sowohl die Freisetzung des Wachstumshormons (GH) als auch die des Dehydroepiandrosterons (DHEA) und des Melatonins nehmen mit der Alterung ab (Jockenhöfel, 1999). Belegt sind veränderte Funktionen der Hypothalamus-Hypophysen-Gonadalachse, der Hypothalamus-Hypophysen-Nebennierenrinden-Achse sowie der Hypothalamus-Hypophysen-Thyroid-Achse (Jones & Boelaert, 2015). Die altersbedingte Abnahme von anabol wirkenden Androgenen führt zur Rückbildung der Muskelfasergröße, zur Zunahme des Bindegewebes und zum Anstieg des Körperfetts. Mit der Abnahme anabol wirkender Hormone (gonadale Steroide) ist eine Abnahme der Muskelkraft vorprogrammiert (Finkelstein et. al., 2013).

Jedoch ist im Spitzensport derzeit zu beobachten, dass noch in der 4. Lebensdekade, wo bereits der Seniorensport beginnt, Höchstleistungen realisiert werden. So lief 6 Monate nach seinem 35. Geburtstag Haile Gebrselassie (ETH) einen Marathonweltrekord in 2:03:59 h in Berlin.
Die Trainingsprinzipien sind für die Seniorensportler, welche niedrigere Trainingsgeschwindigkeiten oder Trainingsumfänge wählen, genauso gültig wie für jüngere Athleten. Das betrifft auch die Belastungsdauer in einer Trainingseinheit. In der Regel trainieren leistungsorientierte Läufer im Seniorenalter bis zur 7. Lebensdekade 4 bis 10 Stunden/Woche. Das Wettkampfresultat in den Ausdauersportarten repräsentiert meist den altersbedingt möglichen Trainingsaufwand (Laufkilometer) und die dabei erreichte mittlere Laufgeschwindigkeit über mehrere Monate vor dem Start (Tanda & Knechtle, 2015). Der nachlassende anabole Stoffwechsel und der Muskelschwund sind im höheren Seniorenalter wesentliche Ursache für die Abnahme von Kraft-, Schnellkraft- und Ausdauerfähigkeiten.
Herausragende Leistungen in den Altersklassen
Die anhaltende Leistungszunahme in den Laufsportarten betrifft nicht nur das Hochleistungsalter, welches in den Ausdauersportarten zwischen dem 25. bis 35. Lebensjahr liegt. Auch die Altersrekorde nehmen gegenwärtig in vielen Sportarten zu. Auffallend ist, dass mit zunehmendem Lebensalter sowohl kürzere als auch längere Distanzen im Visier von Rekordbemühungen sind. Mit 100 Jahren lief 2015 ein amerikanischer Athlet die 100-m in 26,99 s (Tab. 2).
Tab. 2. Auswahl von Rekorden in den höchsten Altersklassen (AK) in der Leichtathletik im Seniorenalter von Männern (Stand 2019).
| Ausgewählte Altersrekorde (Männer) in leichtathletischen Disziplinen, ab AK 80/105 | |||||
| Altersklasse (AK) Männer | Sportart | Leistung | Name (Nation) | Jahr | |
| 100-m-Lauf | |||||
| 90 | 17:53 s | F. Fischel (BRA) | 2007 | ||
| 95 | 20:41 s | F. Fischel (BRA) | 2017 | ||
| 100 | 26:99 s | D. Pellman (USA) | 2015 | ||
| 105 | 34:50 s | S.Kowalski (POL) | 2015 | ||
| Marathon | |||||
| 90 | 6:35:47 h | M.Fremont (USA) | 2012 | ||
| 100 | 8:25:17 h | F. Singh (UK) | 2011 | ||
| Hochsprung | |||||
| 90 | 1,15 m | D. Pellman (USA) | 2005 | ||
| 95 | 1,00 m | E. Zensch (AUS) | 2015 | ||
| 100 | 0,90 m | D. Pellman (USA) | 2015 | ||
| Weitsprung | |||||
| 90 | 3,26 m | D. Pellman (USA) | 2005 | ||
| 95 | 2,15 m | E. Zensch (AUS) | 2015 | ||
| 100 | 1,75 m | D. Pellman (USA) | 2015 | ||
| 10.000-m-Lauf | |||||
| 80 | 42:39:95 min | E. Whitlock (CAN) | 2001 | ||
| 85 | 51:07:05 min | E. Whitlock (CAN) | 2016 | ||
| 90 | 82:21:05 min | A. Althaus (D) | 1994 | ||
Mit 100 Jahren lief der gebürtige Inder F. Singh, jetzt in England lebend, 2011 den Marathon in 8:25:16 h. Der Rekord wurde nicht anerkannt, weil er keine Geburtsurkunde vorweisen kann (Tab. 2).
Sowohl im Hochleistungsbereich als auch im Altersklassensport sind noch keine Leistungsgrenzen erreicht. Im Altersbereich gibt es das Phänomen des Sammelns von Ultraläufen, d. h. alle Distanzen ab Marathonlauf und länger. Der Extremläufer H. Preisler, geb. 1935, schaffte über 1.809 Ultras in offiziell ausgeschriebenen Wettkämpfen. Bei den Frauen hat gegenwärtig die Extremläuferin S. Eichner, geb. 1940, die meisten Ultrawettkämpfe (>2.000) aufzuweisen.
Sie begann mit 41 Jahren mit dem Lauftraining und legte bis Ende 2017 exakt 161.602 km im Training und Wettkampf zurück. In der 7. Lebensdekade nahmen ihre Gesamtbelastung und Marathonlaufzeit deutlich ab, obgleich sie jährlich 3.000 bis 5.000 km lief (Abb. 4). Unabhängig vom Trainingsaufwand, nimmt objektiv die Laufgeschwindigkeit in Seniorensport bei Männern und Frauen ab (Neumann, 2018).
Ein besonderes Beispiel über das Leistungspotential der Senioren ist der Japaner Hiromu Inada. Er nahm erst mit dem 69. Lebensjahr das Triathlontraining auf und beendete mit 79 Jahren den ersten Langtriathlon. Mit 86 Jahre finishte er 2018 in 16:53:50 h den Ironman auf Hawaii (3,8 km S: 1:15:26 h; 180 km Rad (22,3 km/h) 8:02:40 h; Marathon 6:28:18 h).
Tab. 3. Auswahl von Rekorden in den höchsten Altersklassen (AK) in der Leichtathletik im Seniorenalter von Frauen (Stand 2019).
| Ausgewählte Altersrekorde (Frauen) in leichtathletischen Disziplinen, ab AK 75/100 | |||||
| Altersklasse (AK) Frauen | Sportart | Leistung | Name (Nation) | Jahr | |
| 100-m-Lauf | |||||
| 90 | 23:15 s | M. Morita (JAP) | 2013 | ||
| 95 | 30:18 s | D.Friedman (USA | 2017 | ||
| 100 | 39:62 s | J. Hawkins (USA) | 2015 | ||
| Marathon | |||||
| 80 | 4:11:45 h | Y. Nakano (JAP) | 2017 | ||
| 85 | 5:14:26 h | B.J.Mc Hugh (CAN) | 2012 | ||
| 90 | 7:03:17 h | B.J.Mc Hugh (CAN) | 2017 | ||
| Hochsprung | |||||
| 85 | 0,95 m | R. Pedersen (DEN) | 2015 | ||
| 90 | 0,86 m | M. Hinton (USA) | 2011 | ||
| 95 | 0,78 m | O. Kotelko (CAN) | 2014 | ||
| Weitsprung | |||||
| 85 | 2,93 m | R. Pedersen (DEN) | 2015 | ||
| 90 | 1,77 m | O. Kotelko (CAN) | 2009 | ||
| 95 | 1,57 m | O. Kotelko (CAN) | 2014 | ||
| 10.000-m-Lauf | |||||
| 75 | 50:00:93 min | M.Zerwenka-Nagel (D) | 2005 | ||
| 80 | 51:46:65 min | Y. Nakano (JAP) | 2018 | ||
| 85 | 1:26:15:07 min | V.Kokina (UKR) | 2014 | ||
Eine erhöhte sportliche Leistungsfähigkeit befindet sich gegenwärtig noch auf einem allgemeinen theoretischen Erklärungsniveau. Alles im Leistungssport auf das genetische Potenzial zu reduzieren, wäre eine starke Vereinfachung. Neben voraussetzenden Genen werden bereits belastungsreizempfindliche Gene, Gen zu Gen-Interaktionen, Gen-Umwelt-Interaktionen diskutiert.
Die Grenzen der Belastbarkeit sind noch nicht bekannt. Die Deutung der Ursachen von Interaktionen und nicht-genetischen Einflussfaktoren werden unterschieden. Diese Faktoren bilden zusammen die Voraussetzung für Gesundheit und Leistungsfähigkeit des „Sporttreibenden Phänotyps“ (Individuum). Auffallend ist, dass die Rekorde im Seniorensport neuern Datums sind und von Sportlern vieler Nationen erbracht werden (s. Tab. 2 und 3).

Abb. 2 Trainings- und Wettkampfdaten einer Extremläuferin S. E. (geb. 1940) über die Marathondistanz. Die Sportlerin fing erst ab dem 41. Lebensjahr mit dem Laufen an. Die Abnahme der Laufgeschwindigkeit über die Marathondistanz betrug 2,47 %/Jahr.
Zusammenfassung
Die Ursachen der Leistungsabnahme in Ausdauersportarten (Triathlon, Lauf , Skilanglauf u. a.) im Alter sind vielfältig; bedeutsam sind: Sarkopenie, reduzierte Telomerasenaktivität an Chromosomen, Abnahme exprimierter Myokine, verminderte Proteinsyntheseraten, Hormonumstellungen u. a. Faktoren.
Der Geschlechtsdimorphismus der Leistungsfähigkeit gilt auch im Seniorenalter; die Differenz der Geschwindigkeit zwischen Frauen und Männern in den Laufsportarten (Marathon, Halbmarathon, 10.000 m) beträgt ca. 20 % und ist damit größer als im Spitzensport (10-12 %).
Die Leistungsunterschiede im Triathlon zwischen Männern und Frauen sind durch Streckendifferenzen und unterschiedliche Starts nicht genau bestimmbar, sie betragen durchschnittlich 13-15%.
Moderate Ausdauerbelastungen von insgesamt ~ 150 min/Woche (unabhängig von der Sportart) haben den größten Gesundheitseffekt.
Sowohl im Spitzensport als auch im Seniorensport sind gegenwärtig keine Leistungsgrenzen absehbar.
Der Alterssport gehört zu den positiven Aktivitäten, welche den physiologischen Leistungsabbau im Seniorenalter verzögern hilft.
Das Training im Seniorenalter, kann die Abnahme der Laufgeschwindigkeit sowohl im Sprint (100-m-Lauf) als auch in der Langzeitausdauer (Marathonlauf) nicht verhindern. Der Geschwindigkeitsabfall erfolgt bei Männern und Frauen individuell unterschiedlich schnell, er ist bei Frauen meist größer. Der Muskelschwund (Sarkopenie), der bei Trainierenden im Vergleich zu Untrainierten langsamer verläuft, verhindert nicht die Leistungsabnahme, die nach dem 80. Lebensjahr in den leichtathletischen Sportarten besonders deutlich wird. Die Grenze der Belastbarkeit im Seniorenalter ist derzeit bei Männern und Frauen noch nicht erreicht, wie am Beispiel von Extremsportlern zu ersehen ist.

DER AUTOR:
 Prof. Dr. med. habil. Georg Neumann forschte am Institut für Körperkultur und Sport (FKS) Leipzig, arbeitete für das Institut für Angewandte Trainingswissenschaft (IAT) Leipzig im Bereich Leistungssport und für die Universität Halle im Bereich Trainingswissenschaft. Bis zu seinem Ruhestand hat er lange Zeit Leistungssportler im Skilanglauf, Radsport, in der Leichtathletik und zuletzt im Triathlon betreut. Neumann leitete über viele Jahre den wissenschaftlichen Beirat der deutschen Triathlon-Union.
Prof. Dr. med. habil. Georg Neumann forschte am Institut für Körperkultur und Sport (FKS) Leipzig, arbeitete für das Institut für Angewandte Trainingswissenschaft (IAT) Leipzig im Bereich Leistungssport und für die Universität Halle im Bereich Trainingswissenschaft. Bis zu seinem Ruhestand hat er lange Zeit Leistungssportler im Skilanglauf, Radsport, in der Leichtathletik und zuletzt im Triathlon betreut. Neumann leitete über viele Jahre den wissenschaftlichen Beirat der deutschen Triathlon-Union.
Kontakt: neumann.leipzig@t-online.de